Milada Teplá
KUDCH, PřF UK v Praze
email: milada.tepla@natur.cuni.cz
Biochemie - mezioborová témata
- NA: Slovo úvodem
- NA: Studijní text
- NA: Animace
- Trávení: Slovo úvodem
- Trávení: Studijní text
- Trávení: Animace a videa
- Digestion: English version
- Dýchání: Slovo úvodem
- Dýchání: Studijní text
- Dýchání: Animace a videa
- Lipidy: Slovo úvodem
- Lipidy: Studijní text
- Lipidy: Animace a videa
- Signalizace: Slovo úvodem
- Signalizace: Studijní text
- Signalizace: Animace
- Sport: Slovo úvodem
- Sport: Studijní text
- Sport: Výukové materiály
- Volné radikály a Antioxidanty
Nukleové kyseliny, replikace, transkripce, translace a genové inženýrství
Trávicí soustava, trávení přírodních látek
Dýchací soustava, transport kyslíku a oxidu uhličitého a buněčné dýchání (dýchací řetězec a syntéza ATP)
Lipidy, biologické membrány a transport látek přes membrány
Buněčná signalizace
Biochemie a sport
Biochemie v lékařství
Buněčná signalizace - studijní text
Studijní text ke stažení: pdf
Studijní text obsahuje tři kapitoly. První kapitola je kapitolou úvodní, která žáky seznamuje s tématem buněčné signalizace, především s jejím významem. Druhá a třetí kapitola je zaměřena na konkrétní typy přenosu buněčné signalizace. Obsah kapitol odpovídá obsahu výukové animace, i když ne všechny ve studijním textu popisované děje byly zanimovány. Studijní text je doplněn o sedm otázek k procvičení včetně autorského řešení.
Cílem studijního textu je prezentace učiva v podobě textu doprovázeného o statické obrázky, které byly vytvořeny v programu Adobe Animate 2021. Text slouží primárně žákům středních škol a jejich učitelům.
Studijní text byl vytvářen v grafickém editoru Canva.Cílem grafické editace bylo zvýšení atraktivity učiva. Graficky editovaný studijní text byl vytvářen tak, že v hlavním textu žák nalezne základní informace týkající se probíraného tématu, po stranách pak zajímavosti, poznámky či doplňující informace.
*************************************************************************************************************
Buněčná signalizace
Buňka je základní stavební a funkční jednotka těl živých organismů. Aby přežila, má důležitou úlohu, a to komunikaci s ostatními buňkami. Jestliže buňka nedostává signály ke komunikaci, tak není schopná přežít. Během vývoje se buňky dělí a diferencují, což znamená, že každá buňka dostává nějakou funkci. V pozdějším stadiu buňky pomocí komunikace regulují metabolické pochody.
Existuje několik typů buněčné signalizace. Endokrinní signalizace je zprostředkována pomocí hormonů, které se tvoří v endokrinních žlázách a šíří se krví a tkáňovým mokem. Další signalizací je nervová signalizace (synaptický přenos) vyvolaná prostřednictvím nervového systému, která se děje na místě zvaném synapse na konci axonu neuronové buňky. O těchto dvou signalizacích je více řečeno v kapitolách Nervová signalizace – synaptický přenos a Hormony a endokrinní signalizace. Dalšími dvěma typy buněčné signalizace jsou parakrinní signalizace, u které signální molekula difunduje extracelulárním médiem a působí na buňky, které se nacházejí v nejbližším okolí a dotyková signalizace, při které je signalizující a cílová buňka v přímém kontaktu.
Tkáně a orgány lidského organismu jsou koordinovány takzvanými signály, které jsou vyslány signalizující buňkou buňce cílové, která signál přijme, vyhodnotí a odpoví na něj svou reakcí. Signály mohou být chemické látky či fyzikální podněty. Chemickým signálem mohou být hormony (například adrenalin, kortisol), neurotransmitery (například acetylcholin či kyselina γ‑aminomáselná) a další chemické látky. Fyzikálním signálem je elektrický vzruch (akční potenciál, který je vyvolán depolarizací membrány) u synaptického přenosu. Většina signálních molekul z důvodu neprůchodnosti skrze plasmatické membrány nemůže přímo do buňky proniknout. Taková molekula (takzvaný agonista) se následně váže na cílový transmembránový protein – receptor. Nejčastěji se setkáme s třemi typy takovýchto povrchových receptorů. Prvním typem jsou receptory spojené s iontovým kanálem. Druhým typem jsou receptory s vlastní enzymovou aktivitou. Třetím typem jsou receptory spřažené s G-proteiny. Co mají všechny tyto receptory společného je to, že po navázání signální molekuly na vnější straně plasmatické membrány mění svou konformaci (receptor je zaktivován). Tato změna spouští sled událostí (signál se šíří dál), aniž by signální molekula vstoupila do buňky. K odstranění signální molekuly dochází ukončením aktivace receptoru. Ke konci aktivace receptoru dojde, jakmile dochází k informační kaskádě již uvnitř buňky. Buňka odpovídá na signál například regulací genové exprese, regulací metabolické dráhy či změnou cytoskeletu.
1) Synaptický přenos
Synaptický přenos neboli nervová signalizace je jeden z druhů buněčné signalizace. Při této signalizaci je signál, kterým je nervový vzruch, předáván z konce axonu neuronu cílovým buňkám, obvykle dalším neuronům či svalovým buňkám.
Nervové buňky (neurony) se od ostatních buněk velmi liší. Z těla buňky vystupují výběžky zvané dendrity a jeden dlouhý výběžek zvaný axon. Díky dendritům neurony přijímají signály a pomocí axonu je zase vysílají dále. Axony mohou být dlouhé až jeden metr a jsou obklopeny Swannovými buňkami izolovanými lipidovou myelinovou pochvou (obrázek č. 1).
Obrázek č. 1: Popis nervové buňky (neuronu).
V nervových zakončeních axonů jsou synaptické váčky, které obsahují signální látky neurotransmitery. K přenosu signálu dochází na synapsích, tedy místech, ve kterých se dva neurony či neuron a svalová vlákna spojují. Jako synapsi můžeme označit nervová zakončení signalizující (presynaptické) buňky (presynaptická membrána buňky), kde se nachází synaptické váčky s neurotransmitery, povrch cílové(postsynaptické) buňky (postsynaptická membrána buňky), která obsahuje receptory, na které se neurotransmitery váží a úzká synaptická štěrbina mezi těmito buňkami. Odhaduje se, že mozkové neurony jsou v kontaktu s více než tisícem jiných neuronů a že celkově v lidském organismu existuje 1015 synapsí.
Příkladem neurotransmiteru je například molekula acetylcholinu, což je ester kyseliny octové a cholinu (obrázek č. 2).
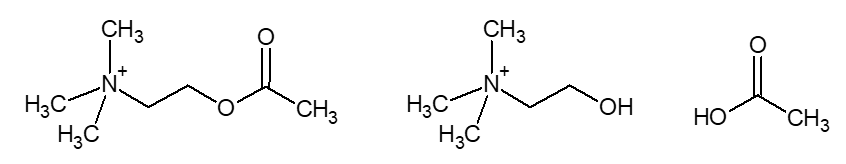
Obrázek č. 2: Vzorec acetylcholinu, cholinu a kyseliny octové.
Acetylcholin účinkuje na nervosvalové ploténce, což je druh synapse mezi neuronem a kosterním svalem. Jeho vyplavení do synaptické štěrbiny způsobuje svalovou kontrakci. Acetylcholin se váže na dva druhy receptorů. První z nich je mimo jiné receptorem též pro alkaloid nikotin (obrázek č. 3) (struktura nikotinového acetylcholinovéhoreceptoru – viz obrázek č. 4), druhý též na alkaloid muskarin (obrázek č. 3), který se vyskytuje například v muchomůrce červené a vyvolává trvalé podráždění svaloviny.
Obrázek č. 3: Struktura alkaloidů nikotinu a muskarinu.
Obrázek č. 4: Model nikotinového acetylcholinového-receptoru, (strukturu lze otevřít na webové stránce pod kódem 6PV7: https://www.rcsb.org/structure/6PV7).
Jak se vlastně acetylcholin objeví v nervovém zakončení axonu? Vzniká už v cytoplasmě presynaptického axonu, a to enzymatickou reakcí acetylkoenzymu A s cholinem. Následně se ukládá do synaptických váčků.
Jak se šíří nervový vzruch?
Signál v podobě akčního potenciálu neboli elektrického vzruchu proudí směrem od těla neuronu axonem k nervovému zakončení signalizující buňky, čímž dochází k lokálnímu vzrůstumembránového potenciálu na membráně presynaptické buňky. Jakmile do nervového zakončení akční potenciál dojde, otevřou se napětím řízené vápenaté kanály, kterými proudí dovnitř signalizující buňky vápenaté ionty. Vápenaté ionty vyvolávají exocytosu synaptických váčků, které obsahují molekuly neurotransmiteru (například acetylcholinu), čímž se elektrický signál převádí na signál chemický. Neurotransmitery se vylučují skrze membránu presynaptické buňky přímo do synaptické štěrbiny, kterou difundují. Následně se vážou na receptory, které jsou spojené s iontovými kanály. Receptory jsou integrovány v membráně postsynaptické buňky.
Díky navázání neurotransmiterů na receptory dochází ke změně jejich konformace a tím k přechodnému otevření iontově sodných kanálů.
Průchod sodných kationtů z extracelulárního prostoru do prostoru intracelulárního vyvolává depolarizaci membrány postsynaptické buňky, čímž se chemický signál mění zpět na signál elektrický, neboť vzniká akční potenciál vyvolávající vzruch, který se šíří dále. Tento děj je znázorněn na následujícím obrázku (obrázek č. 5).
Obrázek č. 5: Nervová signalizace – synaptický přenos.
2) Hormony a endokrinní signalizace
Hormony jsou chemické molekuly s regulačními účinky. Jsou produkovány endokrinními buňkami, které se nachází v endokrinních žlázách (žlázách s vnitřní sekrecí). Od toho název endokrinní signalizace, tedy signalizace iniciována hormony. Hormony mohou být jak lipofilnípovahy (například kortisol, estradiol, testosteron či thyroxin), tak povahy hydrofilní (například adrenalin, glukagon či insulin). Podle povahy hormonu existují dvě odlišné cesty endokrinní signalizace, z nichž každá má zcela odlišný druh receptoru. U menších hydrofilních hormonů musí receptor být uložen v plasmatické membráně cílové buňky, u hydrofobních hormonů leží receptory uvnitř cílové buňky. Hydrofilní hormony se stejně jako lipofilní hormony (a hydrofobní látky obecně) šíří k cílovým orgánům krví nebo tkáňovým mokem, avšak na rozdíl od látek lipofilní (hydrofobní) povahy nepotřebují k tomuto přenosu transportní bílkovinu.
Role receptorů je velmi významná. Například nedostatek receptorů pro mužský pohlavní hormon testosteron, jenž je signálem pro vývoj sekundárních mužských pohlavních znaků, způsobuje, že jedinec se vyvíjí s vnějšími znaky typickými pro ženy (vyvine se jako žena). Příčinou tohoto chybného vývoje tedy není nedostatek hormonu testosteronu, nýbrž nedostatek jeho receptorů.
2A) Endokrinní signalizace – lipofilní hormon
Nejčastějšími lipofilními (hydrofobními) hormony jsou steroidní hormony, mezi které patří například molekula kortisolu (obrázek č. 6), estradiolu či testosteronu (obrázek č. 7). Mezi hydrofobní hormony patří též hormony thyroidní (například thyroxin) (obrázek č. 7).
Obrázek č. 6: Struktura kortisolu – hormon produkovaný kůrou nadledvinek, který pomáhá organismu zvládnout stres či přestát dlouhodobé hladovění.
Obrázek č. 7: Struktura estradiolu, testosteronu a thyroxinu.
Lipofilní hormony jsou transportovány krví pomocí transportní bílkoviny, která je dovede až k cílové buňce. Vzhledem k tomu, že menší lipofilní (hydrofobní) hormony mohou procházet plasmatickou membránou, receptory pro tyto hormony jsou uloženy přímo ve vnitřku buňky, a to buď v cytoplasmě či dokonce přímo v jádře buňky. Receptor, jenž je lokalizován v cytoplasmě, se nazývá cytoplasmový receptor. Jedná se o genově regulační protein, který je bez navázaného hormonu neaktivní. Po navázání steroidního hormonu změní receptor svou konformaci a stane se aktivním. V této aktivní formě se steroidním hormonem vytvoří takzvaný aktivovaný komplex, který je přenesen do jádra skrze jaderné póry a je schopný se v něm navázat na konkrétní sekvenci v deoxyribonukleové kyselině (DNA) a zahájit tak transkripci požadovaného souboru genů. Celý děj je znázorněn na obrázku níže (obrázek č. 8).
Obrázek č. 8: Endokrinní signalizace – lipofilní hormon.
2B) Endokrinní signalizace – hydrofilní hormon
Mezi hydrofilní molekuly, které jsou signálními molekulami v buněčné signalizaci, patří převážně hydrofilní proteiny, peptidy (například insulin, glukagon či antidiuretický hormon vasopresin) a další ve vodě rozpustné molekuly (například adrenalin). 3D struktury některých hormonů jsou znázorněny na obrázcích č. 9 a 10, chemický vzorec adrenalinu je znázorněn na obrázku č. 11.
Obrázek č. 9: Struktura glukagonu – lineární polypeptid, který je tvořen z 29 aminokyselin. Je to hormon produkovaný alfa buňkami slinivky břišní a udržuje v krvi vyrovnanou hladinu glykémie. (strukturu lze otevřít na webové stránce pod kódem 6PHM: https://www.rcsb.org/structure/6PHM)
Obrázek č. 10: Struktura insulinu – skládá se ze dvou polypeptidových řetězců, které jsou spojeny disulfidovými můstky. Dohromady ho tvoří 51 aminokyselin. Je to hormon produkovaný B buňkami Langerhansových ostrůvků slinivky břišní, který snižuje hladinu cukru v krvi. (strukturu lze otevřít na webové stránce pod kódem 2TCI: https://www.rcsb.org/structure/2TCI)
Obrázek č. 11: Struktura adrenalinu – hormon produkovaný dření nadledvinek, který se podílí spolu s kortisolem na udržení organismu při životě během stresové reakce.
Hormony se váží na receptorový protein integrovaný v plasmatické membráně cílové buňky. Nejčastějším typem takovýchto receptorů jsou receptory vázané na G‑proteiny. Tyto receptory mají strukturu polypeptidového řetězce, který sedmkrát prostupuje lipidovou membránou (struktura viz obrázek č. 12) a v neaktivní formě (bez navázané signální molekuly) tvoří komplex s G-proteinem.
Obrázek č. 12: Struktura receptoru vázaného na G-protein. (strukturu lze otevřít na webové stránce pod kódem 4GRV: https://www.rcsb.org/structure/4GRV)
G-proteiny jsou regulační proteiny, které jsou složeny z proteinových podjednotek α, β, γ (obrázek č. 13).
Obrázek č. 13: Struktura neaktivního G-proteinu. (strukturu lze otevřít na webové stránce pod kódem 3AH8: https://www.rcsb.org/structure/3AH8)
V neaktivní formě je na α podjednotku navázán guanosindifosfát (GDP). Po aktivaci receptoru navázáním signální molekuly (například adrenalin či vasopresin), ztratí α podjednotka G-proteinu afinitu ke GDP a získá ji ke guanosintrifosfátu (GTP), což způsobí, že se G-protein uvolní od receptoru a následně se rozpadne na aktivovanou podjednotku α a na komplex β, γ. Dokud je signální molekula na receptoru stále navázána, může receptor aktivovat další G-proteiny. G‑protein, přesněji jeho aktivovaná α podjednotka, se váže na další enzymy (například adenylátcyklasu či fosfolipasu C) či iontové kanály (například iontově specifické vápenaté ionty) a mění tak jejich aktivitu (aktivuje či inaktivuje), což vede k syntéze (nebo naopak k tlumení syntézy) dalších signálních molekul takzvaných druhých poslů (prvním poslem je signální molekula (agonista) vázaná na receptor spjatý s G-proteinem).
Například vazba α podjednotky na adenylátcyklasu vyvolá tvorbu cyklického adenosinmonofosfátu (cAMP) z adenosin-trifosfátu (ATP) a tím zvýší koncentraci cAMP v buňce. Vzniklý druhý posel (cAMP) se v cytoplasmě váže na neaktivní proteinkinasu A (A-kinasu), čímž ji aktivuje. Aktivovaná proteinkinasa A se přesouvá jadernými póry do jádra a následně aktivuje (fosforyluje) či inaktivuje další regulační proteiny (záleží na typu signální molekuly a na druhu cílové buňky). Aktivované fosforylované regulační proteiny jsou schopny se navázat na konkrétní sekvenci v deoxyribonukleové kyselině (DNA) a zahájit tak transkripci požadovaného souboru genů a následnou syntézu nového proteinu (například potřebného hormonu v endokrinních žlázách). I hydrofilní hormon tedy může stejně jako hormon hydrofobní regulovat genovou expresi a s tím spojenou syntézu nových proteinů. Signální dráha, která aktivuje adenylátcyklasu a následně proteinkinasu A je znázorněna na obrázku č. 14. Chemické vzorce GDP, GTP, cAMP a ATP jsou následně zobrazeny na obrázku č. 15.
Obrázek č. 14: Endokrinní signalizace spuštěná vazbou hydrofilního hormonu s aktivací proteinkinasy A.
Obrázek č. 15: Struktury chemických sloučenin: a) guanosindifosfát (GDP), b) guanosintrifosfát (GTP), c) cyklický adenosinmonofosfát (cAMP), d) adenosintrifosfát (ATP).
Druhou častou drahou je vazba α podjednotky na fosfolipasu C, což vede k tvorbě druhých poslů: inositol-1,4,5-trifosfátu (IP3) a diacylglycerolu (DAG) z fosfatidylinositolu (obrázek č. 17), jenž je součástí plasmatické membrány. IP3 způsobí otevření vápenatých kanálů v membráně endoplasmatického retikula, čímž vyplaví vápenaté ionty do cytoplasmy buňky a společně s DAG, jenž je stále součástí plasmatické membrány, následně aktivují proteinkinasu C. Ta se následně podílí na regulaci aktivity nejrůznějších bílkovin (obrázek č. 16).
Obrázek č. 16: Endokrinní signalizace spuštěná vazbou hydrofilního hormonu s aktivací fosfolipasy C.
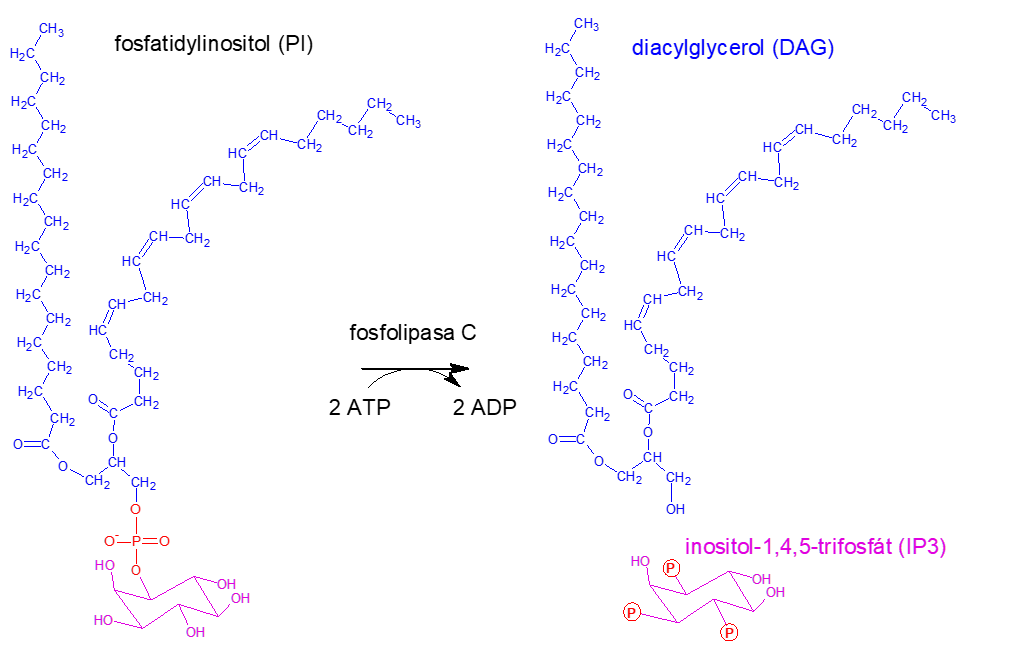
Obrázek č. 17: Tvorba druhých poslů: inositol-1,4,5-trifosfátu (IP3) a diacylglycerolu (DAG) z fosfatidylinositolu (PI).